



Die Geschichte unserer Programmierung: Biologische und soziale Evolution des Homo sapiens
Horden erobern die Erde - Wer sind wir?
Für ein Verständnis der Verhaltensmuster von Homo sapiens - also von jedem Individuum, von menschlichen Rudeln (die heute anders heißen, z. B. Verein, Lobby, Staat) oder von der globalen Spezies als Gesamtheit - kann ein Blick auf die genetische und soziale Programmierung, die uns die Evolution im Verlauf von Millionen Jahren gegeben hat, sicherlich beitragen.
Die folgende Zusammenstellung der biologischen und sozialen Evolution der Gattung Homo bedient sich über weite Strecken zweier Buchquellen, nämlich "Catching Fire" (1) und "The Social Conquest of Earth" (2).
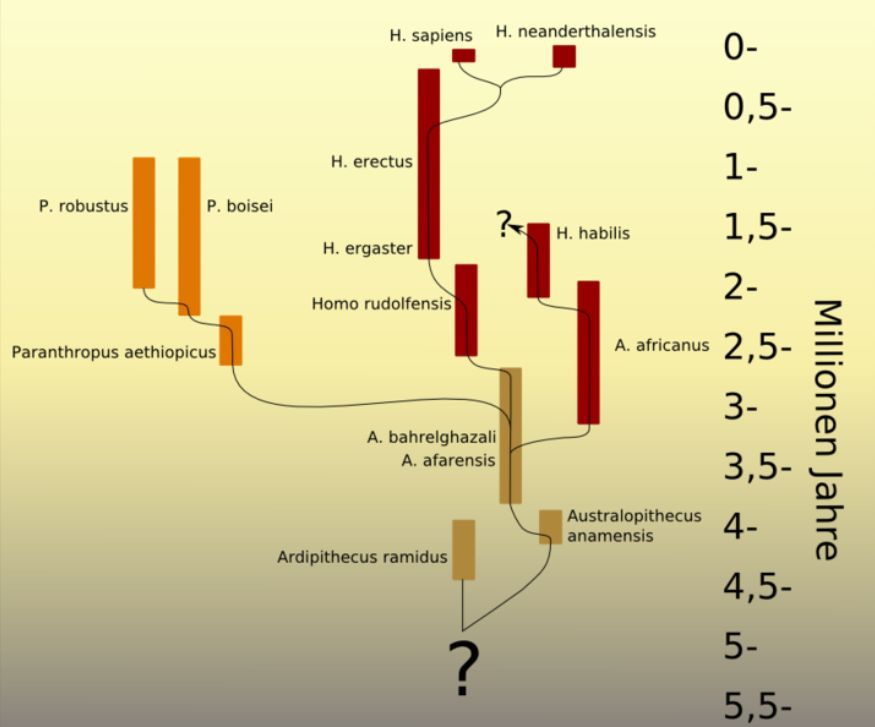
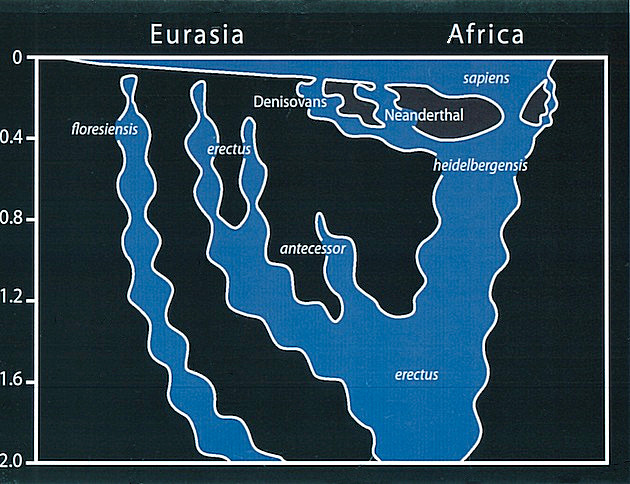
Die folgenden Abbildungen zeigen zwei auf Fossilienfunden und genetischen Analysen beruhende Stammbaumdarstellungen der letzten fünf bzw. zwei Millionen Jahre (Erläuterungen weiter unten).
 |
|
 |
|
| Stammbaumhypothese nach F. Schrenk, dem Erstentdecker (1991) von H. rudolphensis Quelle: Wiki <-- F. Schrenk, "Die Frühzeit des Menschen. Der Weg zu Homo sapiens" (C. H. Beck 1997) |
Stammbaumhypothese nach Ch. Stringer, Mitvertreter der "out-of-Africa"-Theorie, die den europäischen H. neanderthalensis als Seitenlinie und nicht als Vorfahren des H. sapiens identifizierte Quelle: Chris Stringer, Edge.org, 12.11.11 |
Eines der besonderen Merkmale von H. sapiens ist die "Staatenbildung".
Diese wurde von der Evolution schon viel früher und ganz anders realisiert.
Im Paläozoikum, vor ca. 400 Mio. Jahren, entstanden die ersten Insekten, darunter Termiten (220 Mio. Jahre), die Ameisen (150 Mio. Jahre) und die Bienen (100 Mio. Jahre, s. auch Honigbiene: Von der Evolution zur Imkerei). Einige deren Spezies entwickelten die sogenannte "Eusozialität", z. B. die Ameisen vor ca. 100 Mio. Jahren, die Bienen vor ca. 70 Mio. Jahren.
Eusozialität ist ursprünglich definiert über staatenbildende Eigenschaften, nämlich (1.) permanente Gruppen von mehr als einer Generation, (2.) kollektive Brutpflege und sonstige Arbeitsteilung, (3.) Trennung in fruchtbare und unfruchtbare Individuen.
Für den Menschen treffen (1) und (2) zu, und genau betrachtet, in geringem Maß auch (3), z. B. in Ausübung religiöser Regeln (Zölibat) oder, besonders in wohlhabenderen Gesellschaften, in Form von Personen oder Paaren, die aus Karrieregründen bewußt keine Kinder bekommen (wollen). Im Unterschied zu den staatenbildenden Insekten konkurrieren jedoch im allgemeinen bei rudelbildenden Säugetieren die Einzelindividuen in Sexualverhalten und Machtanspruch gegeneinander.
Insbesondere auch unter der evolutionstheoretisch interessanten Thematik "egoistisches vs. altruistisches Verhalten in Gruppen" wird der Begriff "Eusozialität" auch verallgemeinert - den Menschen einschließend - benutzt (2).
Die ersten Säugetiere entstanden im Mesozoikum, vor ca. 150 Mio. Jahren.
Zu Beginn des Känozoikums, vor etwa 55 Mio. Jahren, trennte sich die Ordnung der Primaten (Affen, Menschenaffen, Homo, und deren Vorfahren). Diese besaßen, als Baumbewohner, bereits Greifhände mit ausgeprägter taktiler Sensibilität der Fingerkuppen und flachen Nägeln (statt Klauen), gute Augen und relativ gering ausgeprägten Geruchssinn.
Vor 25 Mio. Jahren trennten sich die Menschenartigen, und daraus vor 8 Mio. Jahren als letzte Aufspaltung der Tribus (~Unterfamilie) der Hominini, der Vorläufer des Menschen, von den Gattungen der Schimpansen und Bonobos, unseren nächsten Verwandten.
Die obenstehenden Graphiken zeigen die weitere Entwicklung, mit Fragezeichen und mit inhaltlichen Unterschieden, die die Ungenauigkeit der Datenlage zeigen. So sind im linken Bild H. floresiensis und H. denisovensis nicht berücksichtigt (letzerer wurde erst 2010 entdeckt), und im rechten Bild ist H. heidelbergensis als Zwischenstufe dargestellt. H. neanderthalensis stirbt aus (links) oder vermischt sich mit H. sapiens (rechts).
In groben Zügen kann gesagt werden, dass ab 4 Mio. Jahren v.u.Z. in Afrika die Gattung Australopithecus nachweisbar ist, ein Hominide mit etwa 450 cm3 Schädelvolumen. Übergangsformen um 2 Mio. Jahre v.u.Z., wie die Spezies H. habilis und H. rudolphensis sind der evolutionären Linie nicht zweifelsfrei zugeordnet. H. erectus, aufrechtgehend und mit einem Schädelvolumen von 900 cm3, ist ab 1,9 Mio. Jahren v.u.Z. nachweisbar.
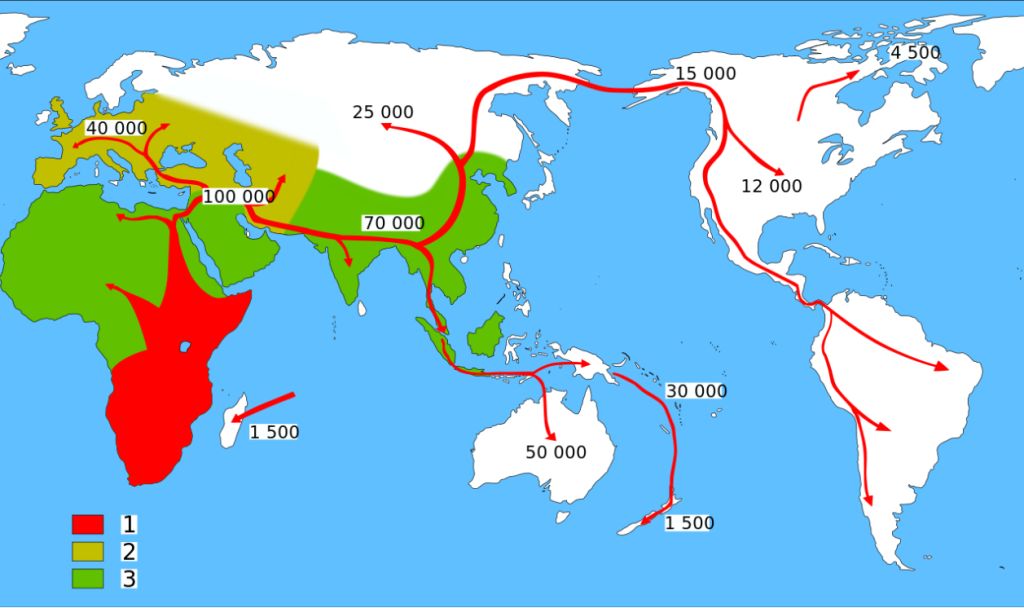 |
|
| Ausbreitung des modernen Menschen (1) über die Erde und vorangehende Besiedelung durch Neanderthaler (2) und andere Hominiden(3) Quelle: Wikipedia, abgerufen 27.11.12 (kein Autor angegeben) |
Er breitete sich, gemeinsam mit später nachgewiesenen Verwandten (600 000 bis
30 000 J. v.u.Z.:
H. neanderthalensis,
H. heidelbergensis, H. denisovensis, H. floresiensis) aus Afrika in Europa und Asien bis China/Indonesien aus.
Die Spezies H. sapiens entstand vor ca. 200 000 Jahren, wiederum in Afrika, und breitete sich im Zeitraum 100 000-1500 J. v.u.Z. global aus, siehe auch die nebenstehende Graphik. Zeitgleich erloschen die Schwesterarten. Kausale Zusammenhänge werden diskutiert, Nachweise sind jedoch schwierig zu führen.
Ein Wort zum Klima: Während des Mesozoikums (vor 250-65 Mio. Jahren) hatte, mit einer Unterbrechung, eine Warmzeit mit etwa 22o Celsius globaler Durchschnittstemperatur geherrscht. Mit Beginn des folgenden Känozoikums (65 Mio. Jahre bis heute) begannen die Temperaturen zu fallen (siehe auch Klimafaktoren der Erdgeschichte).
Vor etwa 30 Mio. Jahren hatte sich Australien so weit von der Antarktis entfernt, dass zirkumpolare Meeresströmungen entstehen konnten, die die Antarktis thermisch abkoppelten und damit deren Vereisung bewirkten und beschleunigten.
Vor etwa 2,5 Mio. Jahren, zu Beginn des Quartärs (2,5 Mio. Jahre bis heute), schloss sich die Meerenge bei Panama. Man diskutiert, dass dies möglicherweise die Meeresströmungen zuungunsten einer Arktis-Heizung veränderte, und auch den Golfstrom in Gang gesetzt haben könnte. Der Nordpolbereich begann ebenfalls zu vereisen. Die globale Durchschnittstemperatur fiel auf 11o Celsius - eine Eiszeit hatte begonnen.
Während dieser Eiszeit schwankten die Temperaturen, verursacht durch Erdbahnoszillationen, im Rhythmus von grob 100 000 Jahren um bis zu ca. 10o Celsius. Die vorletzte Kaltzeit (Glazial) (im Alpenraum: "Riss-Eiszeit") dauerte etwa von 190 000 bis 130 000 Jahren v.u.Z., die letzte (im Alpenraum: "Würm-Eiszeit") von 90 000 bis 10 000 Jahren v.u.Z. Aktuell, im Holozän (10 000 Jahre bis heute), leben wir - innerhalb eines Eiszeitalters - in einer Warmphase.
Auf der oben dargestellten Landkarte erkennt man, dass H. sapiens den Lebensraum Europa, Süd- und Ostasien während des letzten Glazials erobert hat.
Die Warmphase vor ca. 110 000 Jahren bewirkte in Afrika ein trockenes Klima, das möglicherweise die Population von H. sapiens auf einige zehntausend oder weniger Exemplare reduziert und ein Ausweichen nach Norden, durch das Niltal und über die Sinaihalbinsel, begünstigt haben könnte (2). Vor 100 000 Jahren hatte er den "Fruchtbaren Halbmond" erreicht, eine damals klimatisch begünstigte Region vom heutigen Libanon / Syrien über das Zweistromland (Nordirak) bis nach Nordiran. Bis 40 000 v.u.Z. hatte er über das Donautal Europa sowie Asien bis inklusive Australien besiedelt. Zeitgleich starben H. neanderthalensis (Europa), H. denisovensis (Altaigebirge/Sibirien) und H. floresiensis (eine nur einen Meter hohe Zwergform, nachgewiesen in Indonesien) aus.
Die Beringlandbrücke, die durch die starke Vergletscherung um ca. 30 000 bis ca. 15 000 Jahre v.u.Z. geschaffen war (der Meeresspiegel lag mehr als 100 m tiefer als heute), wurde in eben dieser Zeit überschritten und Alaska erreicht Nach Abschmelzen des kanadischen Eisschildes wurde ein Durchgang nach Süden frei, der ab ca. 12 000 v.u.Z. möglicherweise innerhalb eines Jahrtausends die Besiedelung der amerikanischen Kontinente ermöglichte (2; Funde der Clovis-Kultur in Nordamerika bis Mexiko um 11000 v.u.Z.). Die Wanderungsgeschwindigkeit könnte bei über 10 km/Jahr gelegen haben, ein entsprechendes Bevölkerungswachstum vorausgesetzt. Bei einer Vermehrungsrate von 1,3 % pro Jahr (heutige Werte in Entwicklungsländern liegen bei über 3 %) würde sich eine Population von 100 Erstbesiedlern in 1000 Jahren vervierhunderttausendfachen und jeden Quadratkilometer beider Amerikas rechnerisch mit je einem Individuum belegen. Wahrscheinlich gab es in dieser Zeit keinerlei wirksame Predatoren mehr, sodass ausschließlich Nahrungsknappheit bzw. -Angebot die Vermehrungsrate und die Wanderungsgeschwindigkeit bestimmten.
Etwa 1500 v.u.Z. bis 1500 wurden, ausgehend von Tonga, die polynesischen Inseln (im Dreieck Hawai-Osterinsel-Neuseeland) kolonisiert (2).
Die vorgenannten Zahlen sind teilweise nicht unumstritten, für Brasilien werden z. B. auch Zahlen um 30 000 v.u.Z. genannt, ein Szenario, wofür eine Besiedelung über den Pazifik diskutiert wird. Eine Fundstelle in Chile um 14700 v.u.Z. legt nahe, das Prä-Clovis-Menschen schon früher in Amerika waren, sie hätten auch durch Küstenschifffahrt entlang der damals geschlossenen, aber möglicherweise vereisten Beringstraße kommen können.
Die folgende Tabelle stellt nochmals den paläontologischen Zeitstrahl dar (oberer Teil). Im unteren Teil sind einzelne Charakteristika aufgelistet, die nach genetischer Evolution (Körperbau, Gehirngröße u.ä.) und nach sozialer Evolution (von Generation zu Generation gelernte Fähigkeiten und Verhaltensweisen) getrennt sind. Wechselwirkungen sind vorhanden.
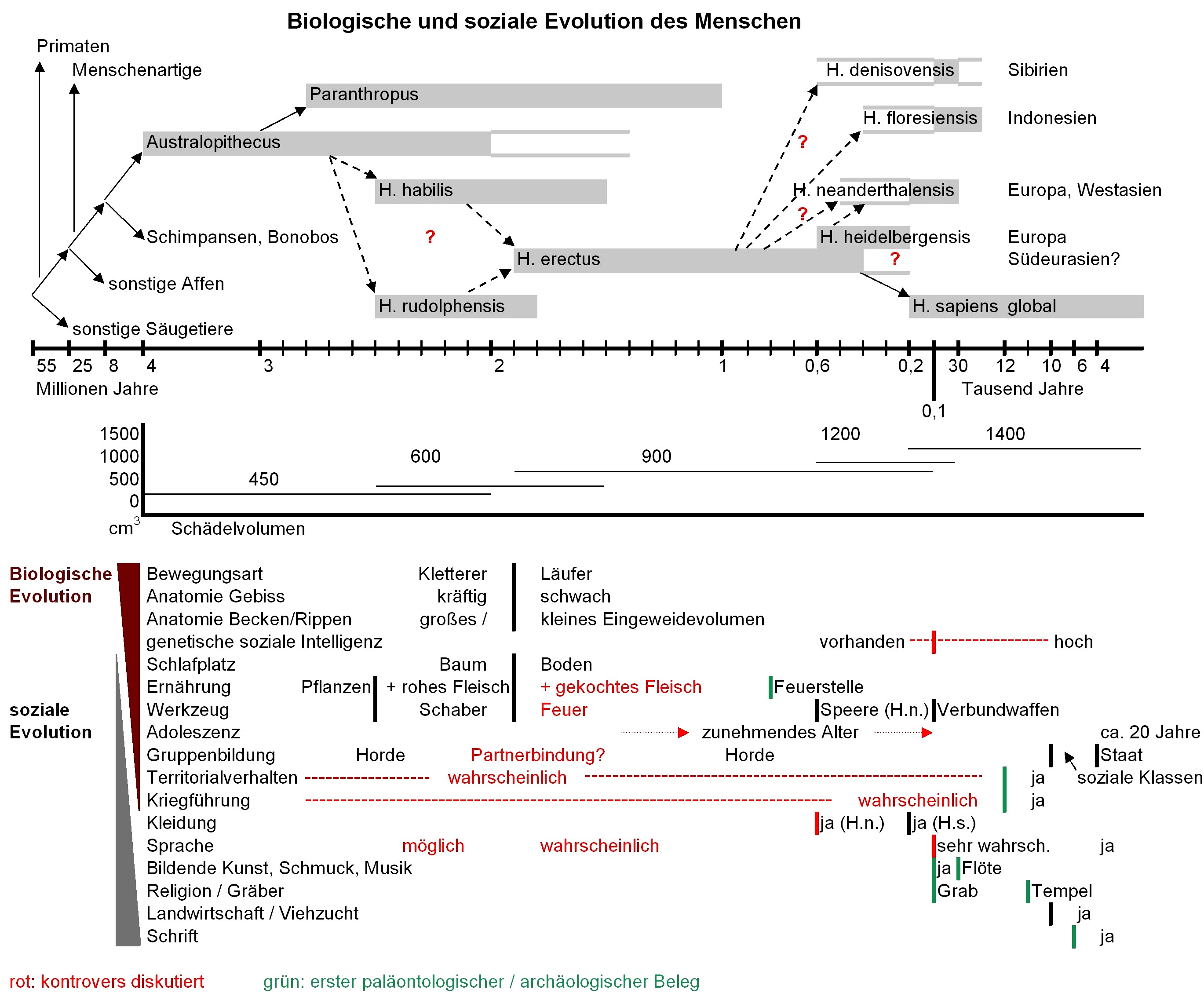 |
||
| zur Beachtung: Die Zeitskalen älter als 3 Mio. und jünger als 0,1 Mio. Jahre sind nicht proportional dargestellt |
Bewegungsart / Anatomie Gebiss-Becken-Rippen / Schlafplatz / Ernährung
Australopithecus war den heutigen Menschenaffen sehr ähnlich. Er war Baumbewohner, mit entsprechend ausgebildeter Schulter- und Armskelett-Ausprägung und Greifhänden, wohl reiner Pflanzenfresser, abzuleiten aus Kiefer- und Zahnformen sowie der relativen Größe von Rippenraum und Becken, aus denen das Volumen der Eingeweide (Magen, Dünndarm, Dickdarm) abgeleitet werden kann. Seine Gehirngröße entsprach ebenfalls heutigen Menschenaffenarten (vgl. Gorilla 550 cm3, Schimpanse 400 cm3). Da die Flora und Großfauna in seinem Habitat, Afrika, funktionell ungefähr den heutigen Savannenlandschaften entsprach (eine Mischung von Flußläufen, Hochgrasflächen und eher offenen Baumbeständen, mit grasfressenden Säugetieren und deren Predatoren, wie z. B. den heute ausgestorbenen Säbelzahntigern [u.a. Gattung Megantereon] oder den jaguarähnlichen Vertretern der Gattung Dinofelis), wird Australopithecus wie seine heutigen Nachfahren in den Bäumen geschlafen haben (2).
Vor ca. 2,8 Mio. Jahren entwickelte sich als Seitenlinie Paranthropus, mit einem kräftigeren Gebiss für härtere Pflanzennahrung (Körner, Früchte), vermutlich wegen einer klimatischen Veränderung (Trockenheit).
Die Zeit vor 2,5-2,0 Mio. Jahren ist fossil nur gering belegt. H. habilis wies als wesentlichen Unterschied ein deutlich größeres Gehirn auf (650 statt 400 cm3). Geringfügige Veränderungen am Gebiss lassen auf einen kleinen Fleischanteil schließen (3). Die ersten Feuersteinabschläge (schmale Bruchstücke mit scharfen Kanten aus dem SiO2-reichen, isotropen Mineral Flint, auch Feuerstein genannt) und Schab- oder Schnittmarken an fossilen Tierknochen deuten darauf hin, dass Fleischteile von Aas gegessen wurden (1).
Der seit 1,9 Mio. Jahren nachweisbare H. erectus war in Bezug auf sein Skelett in mehrfacher Hinsicht verändert. Neben einem wiederum größeren Gehirn (900 cm3) sind Brust- und Beckenraum kleiner, d. h. er hatte einen kleineren Verdauungstrakt. Er ging aufrecht und hatte seine Kletterfähigkeit weitgehend verloren. Gebissstärke und Zahnfläche waren reduziert, also untypisch sowohl für einen reinen Pflanzenfresser als auch für einen reinen (Roh-)Fleischfresser.
Wissenschaftler sehen als einfache und bisher einzig mögliche Erklärung für die Gesamtheit dieser Veränderungen die Beherrschung des Feuers und des Kochens (die älteste nachgewiesene Feuerstelle ist 1,1 Mio. Jahre jünger und datiert von 790 000 v.u.Z., in Israel, wo man verbrannte Samen, Holz und Flintstein fand).
Die "Expensive Tissue Hypothesis" (Hypothese des aufwändigen Körpergewebes) stellt fest, dass der körpergrößenbezogene kalorische Grundumsatz bei Säugetieren unabhängig von der Gehirngröße ist. Gehirngewebe und auch Darmgewebe hat einen hohen Energiebedarf. Die Hypothese lautet, dass in der Evolution Gehirnwachstum stets mit der Reduzierung der Eingeweidegröße einherging, und dass dies jeweils nur mit einer Aufwertung der Nahrung erfolgen konnte (4).
Die "Cooking Hypothesis" sagt, dass diese Aufwertung nur mit Kochen (sowie zusätzlich Zerkleinern) der Nahrung möglich war. Quellung und Verkleisterung von Stärke in pflanzlicher Nahrung sowie Gelatinierung von Kollagen und Denaturierung von Eiweiß in Fleisch, beides chemische Prozesse, die etwa oberhalb 70o Celsius ablaufen, erhöhen die kalorische Ausbeute im Dünndarm durchschnittlich um 20-30% (1). Gleichzeitig verringerte sich die pro Tag mit Kauen verbrachte Zeit von ca. 6 h (Schimpanse) auf ca. 1-2 h (Mensch). Erst dadurch waren mehrstündige Intervalle frei, die z. B. für die reguläre Jagd genutzt werden konnten (1).
Die reguläre (tägliche) Kochpraxis legt die Existenz permanenter Feuerstellen nahe, die damit auch den Schlafplatz am Boden (anatomisch bedingt nicht anders möglich) erklärt. Säbelzahntiger waren von einer Feuerstelle einfach fernzuhalten.
H. erectus war die erste Menschenart, die sich aus Afrika heraus verbreitete. Er ist in Asien/Indonesien seit ca. 1,6 Mio. Jahren, in Europa (Spanien) seit 1,4 Mio. Jahren nachweisbar (1). Die Beherrschung des Feuers könnte sein Ausbreiten von tropischen in gemäßigte Klimata erklären helfen.
Die "Koch-Hypothese" ist mangels direkter Belege nicht unumstritten.
Steinzeit-Feuerzeug: Flint (das allgemein benutzte Werkzeugmineral, das eine hohe Härte hat) wird gegen Pyrit (FeS2, ebenfalls ein häufiges Mineral) geschlagen und löst unter Wärmebildung daraus kleine Partikel ab. Diese verbrennen exotherm (zu Fe2O3 und SO2) unter Flammenbildung. Damit wird Zunderschwamm (z. B. aus dem gleichnamigen Totholz-Pilz) angezündet. Zunderschwamm wurde bei "Ötzi", der Gletschermumie aus den Ötztaler Alpen / Österreich (3000 Jahre v.u.Z.) gefunden.
Seit grob 0,5 Mio. Jahren sind H. heidelbergensis und H. neanderthalensis nachweisbar, in Europa und Vorderasien. Ihrem wiederum größeren Gehirn (1200 cm3) entspricht eine verbesserte Waffentechnik. Auf dem wahrscheinlich zweitältesten Feuerstellenfund (400 000 Jahre v.u.Z., H. heidelbergensis) in Schöningen/Deutschland wurden Holzspeere gefunden, neben Pferde-, Bison-, Esel- und Hirschknochen. Ab ca. 60 000 Jahren v.u.Z. sind zahlreiche Feuerstellen von H. neanderthalensis belegt. Eine interessante Frage ist, warum sich diese beiden Arten im Gegensatz zum vorangehenden H. erectus und zum nachfolgenden H. sapiens geographisch nicht weiterverbreiteten - und warum sie quasi gestern (H. neanderthalensis vor 30 000 Jahren) ausstarben.
Werkzeug
 |
|
| Clovis-Speerspitzen aus der "Rummells-Maske"-Fundstelle, Iowa, USA, ca. 11 000 J.v.u.Z. Quelle: Wikipedia <-- B. Whittaker . |
Zu den Faustkeilen steigender Qualität (H. habilis, H. erectus) und den Äxten, Stein-Speerspitzen und Holzspeeren (H. heidelbergensis, H. neanderthalensis) kamen vor ca. 70 000 Jahren mit H. sapiens Waffen in Verbundbauweise hinzu, z. B. Speere, bei denen die Steinspitze mit einer Mischung aus Baumharz und Bienenwachs eingeklebt war (2). Deren Komplexität - verschiedene Materialien einer Temperaturbehandlung zu unterziehen - setzte eine höhere kognitive Leistung voraus.
Die Abbildung zeigt Speerspitzen der Clovis-Kultur (Nord- und Südamerika, um 11 000 J.v.u.Z.). Diesen Speerspitzen standen 34 Gattungen von Säugetieren mit einem Gewicht über 50 kg in Nordamerika, und 51 Gattungen in Südamerika (11 davon gemeinsam) entgegen, die das folgende Jahrtausend nicht überlebten. Gleichzeitig allerdings erhöhte sich mit dem Ende des letzten Glazials die globale Durchschnittstemperatur innerhalb von 7000 Jahren um etwa 7o Celsius. Die Ursachenzuordnung zu dem Massensterben von Großsäugern ist daher umstritten (11).
Genetische soziale Intelligenz / Adoleszenz
Eine "Kultur" - im Sinne eines unterschiedlichen Verhaltens voneinander getrennt lebender Gruppen derselben Spezies durch Lernen innerhalb dieser Gruppe - haben z. B. Schimpansen (manche Populationen können Steine werden, mit Stöcken Termiten aus ihren Bauten "angeln", kleine Tiere jagen usw., andere nicht). Dazu sind kognitive Fähigkeiten, wie Langzeitgedächtnis, Imitationsfähigkeit, Abstraktionsvermögen und - auch zukunftsgerichtete - Szenarienbildung ("wenn ich den Stock in den Termitenhaufen stecke, werden sich die Termiten dran festbeißen, sodass ich sie herausziehen und essen kann") erforderlich.
Für ein "soziales" (in Gruppen lebendes) Tier liegt es im "persönlichen" Interesse, die Intentionen anderer Gruppenmitglieder zu erkennen und danach geeignet zu handeln, um Vorteile bezüglich Futter, Hierarchie, Sexualpartner usw. zu erlangen. Dem gegenüber steht das Gruppeninteresse, z. B. gemeinsam zu jagen oder den Schlafplatz zu verteidigen. Sofern die Evolution die Gruppe als überlebensfähig gezeigt hat (andernfalls wäre das Einzeldasein vorteilhafter), bedeutet dies automatisch und systemimmanent einen dauerhaften Interessenkonflikt zwischen "Egoismus" und (gruppenbezogenem) "Altruismus".
Die "Social Brain Hypothesis" (5) unterstellt nun, dass die Großhirnentwicklung von Primaten weniger durch die Verarbeitung externer Eindrücke, sondern durch die Optimierung sozialer Verhaltensweisen in Gruppen (zunehmender Größe) evolutionär beeinflusst wird.
So wird insbesonere auch für H. sapiens eine Mutation oder Mutationsserie vermutet, die nicht nur das Großhirnvolumen, sondern auch dessen Funktionalität veränderte, vereinfacht gesagt in Richtung besserer Verschaltung der verschiedenen Gehirnareale.
Als starker Hinweis hierauf gilt der Ausbreitungserfolg von H. sapiens gegenüber seinen Vorläufern ("der Neanderthaler hatte diesselben Chancen") und die ausschließlich auf diese Spezies begrenzte weitere kulturelle Entwicklung (s. weiter unten) (2).
Die - durch ihre evolutionäre Selektionsrelevanz definierte - soziale Intelligenz kann folgendermaßen formuliert werden (frei nach (2)):
- Was tut der andere: Geteilte Wahrnehmung (das Individuum achtet auf das Verhalten anderer Gruppenmitglieder)
- Was will der andere: Erkennen der Intentionen der Gruppenmitglieder
- Was will ich: Szenarienbildung unter Einschluss der eigenen Person (mentale Abbildungen vergangener und zukünftiger eigener und fremder Handlungsalternativen und deren Bewertung für den eigenen Erfolg)
Der letzte Punkt kann als Formulierung für den Begriff "Bewußtsein" verstanden werden.
Informationstechnologisch formuliert, ist die "Hordenwesensoftware", also die Eigenschaftskombination Egoismus / Gruppenaltruismus, innerhalb von mindestens 8 Mio. Jahren eingeführt und in Schritten - parallel mit der Hardware der vererbten Gehirncharakteristika - weiterentwickelt worden, wogegen die Softwareergänzung "Bewußtsein" erst vor einigen zehntausend Jahren zur Blüte kam. Zur Verschaltung insbesondere der sozial relevanten älteren und jüngeren Funktionen (emotionales vs. "rationales" Urteilen) siehe z. B. die "Theorie der somatischen Marker".
Dem erhöhten Lernaufwand entspricht eine verlängerte Kindheits- und Jugendphase. Schimpansen haben eine Adoleszenzphase von ca.13 Jahren, Menschen von ca. 20 Jahren.
Gruppenbildung / Territorialverhalten
Die entfernter verwandten Gibbons ("kleine" Menschenaffen) leben monogam in Territorien. Das Terrtiorialverhalten von Gorillas und Orang-Utans, sogenannten "großen" Menschenaffen, ist gering ausgeprägt. Sie leben polygam, genauswo wie unser nächsten Verwandten, die Schimpansen und Bonobos.
Aus deren mit H. sapiens gemeinsamen ebenfalls starken Territorialverhalten kann geschlossen werden, dass es auf den gemeinsamen Vorfahren, also bis zu 8 Mio. Jahre, zurückgeht.
Die Funktionalität der Gruppe dürfte sich, wie oben bereits geschildert, mit den Meilensteinen "Schlafplatz um Feuerstelle" (H. erectus) und "gemeinsame Jagd" intensiviert haben. Eine Theorie besagt, dass sich die Paarbildung (soziale Monogamie) im Verlauf des Meilensteins "Kochen" entwickelt haben könnte, analog etwa der Kultur heutiger Ureinwohner, bei denen teilweise die Männer tagsüber jagen und die Frauen Früchte und Knollen sammeln, um am Nachmittag für den heimkehrenden Partner zu kochen (2). Die Arbeitsteilung war geboren.
Spätestens zeitgleich mit der Erfindung von Ackerbau und Viehzucht ab etwa 7000 v.u.Z. (s. weiter unten) bildeten sich aus den Gruppen mit persönlich erworbenen Rangordnungen Stammestümer mit mindestens zwei erblich oder durch Rituale festgelegten Hierarchiebesetzungen. Staaten entstanden, ebenfalls unabhängig voneinander, etwa 3400 v.u.Z. (Ägypten), 2900 v.u.Z. (Indien) oder 1800 v.u.Z. (China), um nur einige zu nennen.
Die politisch organisierten Machtgebilde wurden immer größer, z. B. Chinesische Reiche 1000 v.u.Z. bis 19. Jht., (West-)römisches Reich 1000 v.u.Z. bis 500, Mongolisches Reiterhordenreich 1200-1400, koloniales Spanisches Reich 1500-1600, Britisches Empire 1800-1900, USA / Sowietunion im kalten Krieg 20. Jht., globale Spieler (USA, China, Europäische Union und andere) heute. Transnationale Interessenverbände kamen hinzu (Vereinte Nationen 1945, Militärpakte, Wirtschafts- und Handelszonen, Finanzlobby, Klima- und Naturschutzlobby u.ä.).
Der Treiber war stets der Machtanspruch auf Ressourcen. Die Entwicklung lief von reinen Territorialreichen zu politischen, wirtschaftlichen und ideologischen Einflusszonen und hat in einigen Fällen die globale Dimension erreicht (als erstes plakatives Beispiel kann die vertragliche Aufteilung der Erde zwischen Spanien und Portugal im Vertrag von Saragossa 1529 gelten).
Dies alles begann mit einer Horde Hominiden. Wir waren als Hordenwesen erfolgreich gegen Prädatoren, gegenüber dem Neanderthaler, gegenüber widrigen Umwelteinflüssen. Nun sind wir so erfolgreich im Kampf um Ressourcen, dass wir deren Degradation und Zerstörung auslösen.
Die gängige Hypothese lautet, dass die gesellschaftliche Entwicklung der letzten zehntausend Jahre (oder länger) durch keinerlei genetische Veränderungen, sondern ausschließlich durch die soziale Evolution getrieben wurde.
Kriegführung
Auch hierbei nutzt man den Vergleich mit nächsten Verwandten. Schimpansen führen territoriale Eroberungskriege, indem männliche Banden sich in fremdes Territorium schleichen, um einzelne gegnerische Individuen zu suchen. Männliche erwachsene Gegner sowie Junge werden üblicherweise getötet, Weibchen in Ruhe gelassen. Zieht die unterlegene Gruppe irgendwann ab, ist deren Territorium gewonnen (2,6).
Paläontologische Untersuchungen über Knochenschädigungen durch Waffeneinwirkung, aus Fundstellen ab 12 000 v.u.Z. in verschiedenen Kontinenten, ergeben eine Sterberate durch Kriegführung zwischen 0 und 46 %, mit einem Mittelwert um 15 %. Vergleichszahlen von heute, aus Stämmen in Südamerika, Australien und den Philippinen, liefern Prozentsätze zwischen 5 und 30 % (Datenzusammenstellung durch Bowles, (7)).
Daraus kann abgeleitet werden, dass territoriale Kriegführung mit Tötung des Gegners von Entstehung des H. sapiens an, möglicherweise schon vorher seit 8 Mio. Jahren, genetisch und sozial angelegt ist. In der Phase der abstrakten Bewußtwerdung (siehe unten "Kleidung/Sprache") vor grob 100 000 Jahren bildeten sich daraus Begriffe wie "Ehre", "Feigheit", "Treue", "Verrat". Krieg ist eine Ausprägung des Hordeninstinktes von H. sapiens im Konkurrenzkampf der Horden untereinander.
Bowles (7) geht sogar weiter und stellt die Frage, ob die permanente Kriegführung der Horden während mindestens vieler zehntausender Jahre nicht sogar einen Evolutionsdruck in Richtung Intra-Gruppen-Altruismus ausgeübt haben könnte. Eine Horde von "Altruisten" - die sich für den Hordensieg zu opfern bereit sind - würde eine Horde von "Egoisten" durch bessere Taktik besiegen (Waffengleichheit vorausgesetzt), und damit würden sich die "altruistischen Horden" erfolgreicher ausbreiten. Ob die Intra-Gruppen-Altruismusfähigkeit genetisch oder sozial weitergegeben wird, spielt in diesem Zusammenhang keine wesentliche Rolle.
Die "Erfolgsgeschichte" der Besiedelung fast der gesamten Erde durch H. sapiens in wenigen Jahrzehntausenden war möglich einerseits durch Abwesenheit wirksamer Prädatoren und Anpassung an diverse klimatische Habitate, andererseits durch überaus erfolgreiche Nahrungsbeschaffung im Hordenkollektiv in verteidigten oder neuen Territorien: Krieg führen oder wandern.
Kleidung / Sprache
Man vermutet, das H. neanderthalensis Kleidung trug; so fand man in Frankreich (Arcy sur Cure) Ahlen mit deutlichen Gebrauchsspuren, die zum Nähen gedient haben könnten (35 000 J. v.u.Z.); sichere Nachweise gibt es jedoch erst für H. sapiens.
Hinweise auf Sprache sind naturgemäß indirekt. Bereits H. habilis könnte eine Sprachfähigkeit besessen haben, wie aus Schädelformen, die "Brocas Areal" und "Wernickes Areal" (für die Sprache notwendige Gehirnbereiche, s. auch "Somatische Marker") ausgeprägt erscheinen lassen, geschlossen werden kann.
Eine andere Hypothese kommt aus der sogenannten "kognitiven Archäologie". Einer gewissen technischen Komplexitätsfähigkeit, in diesem Fall z. B. für die Herstellung von Verbundwerkstoffen (Mehrkomponentenkleben von Waffen, s. o.) wird eine analoge begriffliche Komplexitätsfähigkeit zur Bildung von Worten und sprachlicher Syntax zugeordnet. Vertreter dieser Theorie schätzen damit ein Alter der Sprache auf nicht unter 70 000 Jahren (8).
Bildende Kunst / Religion
Vor ca. 80 000 Jahren datieren die ersten Schmuckfunde, durchbohrte Schneckenschalen, sowie eine abstrakte Zeichnung (Striche und Punkte) aus Ocker. Zahlreiche Höhlenmalereien von H. sapiens ab etwa 40 000 Jahren v.u.Z. wurden in Europa, schwerpunktmäßig im Südwesten (zur Erinnerung: Die Würmeiszeit war im Aufbau begriffen) gefunden: Chauvet (Frankreich) ab 37 000 Jahren, Altamira (Spanien) ab 22 000 Jahren, Lascaux (Frankreich) ab 17 000 Jahren. Auch anderswo wurde die Felsmalerei erfunden, z. B. in der Serra da Capivara (Brasilien) ab 10 00 Jahren v.u.Z., s. Fotoreihe.
 |
 |
 |
 |
 |
 |
 |
 |
| Serra da Capivara, Piauí, Brasilien, 2007; s. auch Diashow "Serra da Capivara" (13 min) |
|||||||
Musik ist erstmals nachweisbar über Flöten aus Vogelknochen mit einem Alter von ca. 30 000 Jahren. Aus verhaltenspsychologischen Überlegungen heraus wird jedoch die Meinung vertreten, dass Musik, z. B. über den Rhythmus (Trommelschläge u.ä.), Tanz und intonierte Sprache, parallel zu dieser entstanden sein könnte. Hinweise dafür sind einerseits die Analogbeispiele heutiger Eingeborenenkulturen, die das Auftreten von Gesängen vor der Sprachdifferenzierung zeigen, andererseits die gruppenmotivierende Funktion von gemeinsam vollzogenen Riten, darunter solche musikalischer Natur (9).
Die ältesten Gräber mit Hinweisen auf Rituale, an Grabbeigaben zu erkennen, sind etwa 95 000 Jahre alt (Israel). Eine tempelartige Gebäudestruktur, mit Tier- und "Fabel"-Wesenreliefs in der Türkei wurde auf etwa 11 000 Jahre v.u.Z. datiert; ihr Bau fiel möglicherweise mit den Anfängen des Ackerbaus im "Fruchtbaren Halbmond" zusammen (s. unten).
Ab einer gewissen Bewußtseinsstufe - die Datenlage deutet erst auf H. sapiens - wird sich der Mensch gefragt haben, wo er herkommt, wer er ist, und wo er nach dem Tode hingeht. Träume und drogeninduzierte Zustände (bei heutigen Ureinwohnern und in der jüngeren Geschichte zahlreich belegt) boten Bilder und subjektive Erfahrungen, die als Visionen einer äußeren Wirklichkeit gedeutet werden konnten.
Evolutionär wird Stammesriten und -dogmen, darunter Schöpfungsmythen, Identifikationsankern und Heilsversprechungen, eine gruppenstärkende Funktion zugeordnet. Auch um diese "spirituelle Ressource" fanden Kämpfe zwischen den Gruppen statt. So vereinnahmten die monotheistischen Religionen konkurrierende Gottheiten (Judentum ab ca. 1000 v.u.Z., Christentum im Römischen Reich ab ca. 300), Religionen lösten Kriege aus (Islam ab 600, Christentum 1000-1300 [Kreuzzüge]) oder versuchten ihre politische und wirtschaftliche Macht zu erweitern (Buddhismus in China 600-900, jesuitische Christen in Japan 1600-1700) (Jahreszahlen grob gerundet). Nicht-transzendentale Dogmensysteme kamen hinzu (Sklaverei in der westlichen Welt 16. bis 19. Jht., Nationalsozialismus, stalinistischer Kommunismus, Maoismus 20. Jht.).
Religionen bzw. Wertesysteme bieten dem sozialen Instinkt des Hordenmitglieds Geborgenheit, Empathie, Normen und Kontrollmechanismen, fördern die Kohäsion der Gruppe und können hohe innere Energien freisetzen.
Gängige Religionen und Ideologien beinhalten tabuartige Grundregeln, die von Mitgliedern nicht hinterfragt werden dürfen, sowie - evolutionskonform - eine Abgrenzung zwischen Gruppenmitgliedern und -Nichtmitgliedern. Ein Überschreiten der Grenze ist nur nach Anerkennung der Glaubensgrundregeln zulässig.
Der heutige Stand des Wissens über die äußere Welt und unsere Wirkung auf selbige, unter Berücksichtigung der Begrenztheit der globalen Ressourcen und unter gedanklicher Einbeziehung unserer Nachfahren in die eigene "Horde", lässt es zielführend erscheinen, eine Ideologie mit der Grundregel "Wir wollen unsere Zukunft sichern" zu formulieren. Die daraus abzuleitenden Verhaltensregeln betreffen die Spezies als solche.
Das bedeutet, es gibt - evolutionär erstmalig - keine Nicht-Gruppenmitglieder, also keine äußeren Konkurrenten mehr.
Die Identifikation mit der gesamten Spezies als eigener "Horde" umgeht die evolutionäre Prägung der dogmatischen Überheblichkeit der "eigenen" Ideologie gegenüber Außenstehenden. Kennzeichen hierfür wäre eine offene und systemkritische Diskussionsbereitschaft mit "Desinteressierten". Einige heutige Wertesysteme, meist globaler Natur, zeigen diesbezüglich Ansätze: Menschenrechte, Demokratieverständnis, wirtschaftliche Fairness, Nachhaltigkeit (Wirtschaft, Klima, Naturschutz, Bevölkerungskontrolle).
Landwirtschaft / Viehzucht
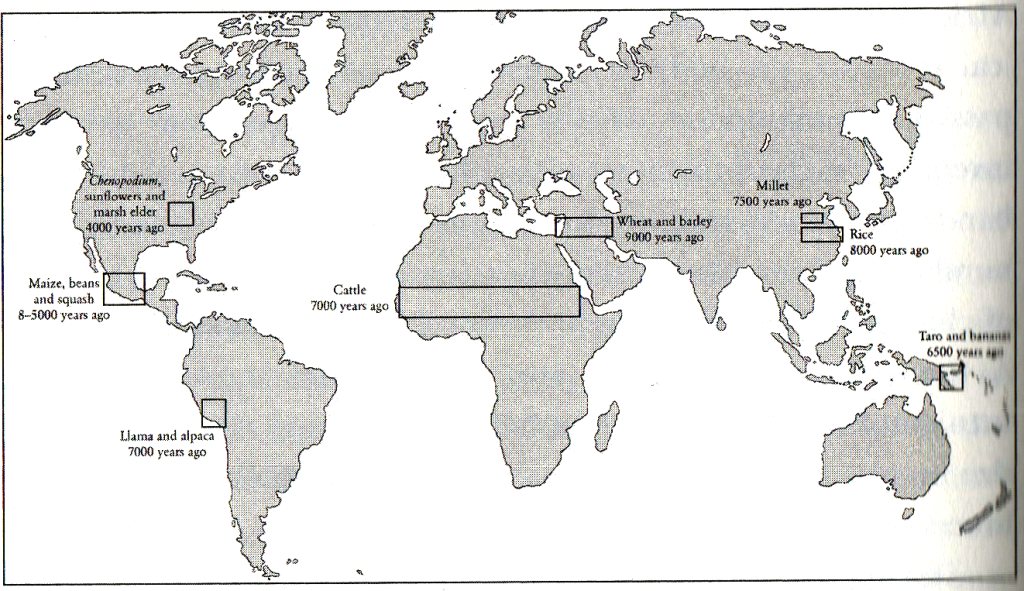 |
|
| Die acht bekannten Regionen der Erfindung des Ackerbaus, inklusive Tierzucht, mit Daten ihrer Entstehung Quelle: S. Mithen, "Did farming arise from a misapplication of social intelligence?", Philosophical Transactions of the Royal Society B 362: 705-718 (2007) |
Mit der Erfindung des Ackerbaus 7 000 bis 2000 v.u.Z. an mindestens acht unterschiedlichen Orten, vermutlich unabhängig voneinander, wurde die Nahrungs-Ressourcennutzung auf eine neue Stufe gehoben, was sich in beschleunigtem Bevölkerungswachstum ausdrückte. Der verringerte Zeitaufwand zur Nahrungsbeschaffung schaffte Freiräume für weitere Tätigkeiten und komplexere Organisationsformen (Architektur, Handwerk, Handel usw., Staatenbildung s. o.).
Nach einer Überschlagsrechnung stieg die Wachstumsrate der globalen Population von 0,013 % pro Jahr (Altsteinzeit) auf 0,05 % pro Jahr (Neusteinzeit bis 1900). Im zwanzigsten Jahrhundert beschleunigte sich das Wachstum weiter auf durchschnittlich 1,3 % pro Jahr (10). Nach einem Maximum um 1960 von 2,5 % fiel der Wert auf heute 1,5 % (Stand 2011).
Es folgen zwei Zitate aus "The Social Conquest of Earth" von E. O. Wilson (2) (Übersetzung G.Mair):
"Es gab keine Möglichkeit, die Konsequenzen unseres anfänglichen Erfolges [Besiedelung fast der gesamten Erde und Erfindung des Ackerbaus] vorherzusehen. Wir führten enfach aus, was uns gegeben war, und fuhren fort, uns zu vermehren und zu konsumieren, in blindem Gehorsam zu Instinkten, die wir von unseren simpleren, stärker in Schach gehaltenen paläolitischen Vorfahren geerbt hatten."
"Ein entscheidender Wandel - bedeutend letztlich nicht nur für die Menschheit, sondern auch für die übrigen Lebewesen, waren die neuen Wertvorstellungen über die Umwelt, die sich in den Köpfen der neugebackenen Bauern bildeten... Natürliches Habitat wurde zu Landfläche, die zum Roden für den Ackerbau bereitstand. Diese spezielle Konzeption, dass Natur zum Ersetzen da sei, ist eine geistige Fixierung der meisten heutigen Menschen."
Schrift
Älteste Nachweise finden sich in Mesopotamien (heute Irak) und Ägpyten um 4400 v.u.Z., in der Induskultur (heute Pakistan) um 2500 v.u.Z. und in China um 1500 v.u.Z. (s. auch Shang-Dynastie in "Geschichte Chinas").
Die Erfindung des Laut-Alphabetes und der dekadischen "arabischen" Zahlen folgten.
Buchdruck (Blockdruck in China 9. Jht., Letterndruck in Europa 15. Jht.) und elektronische Informationstechnologie (20. Jht.) erlaubten die Speicherung und Verbreitung immer größerer Datenmengen.
Die Erzeugung von Wissen und Meinungen und deren Verbreitung ist Katalysator, ist Risiko und Nutzen zugleich für die soziale Evolution des Hordentieres H. sapiens auf globaler Ebene (s. auch Zivilisation 2.0 / Aktionsmöglichkeiten).
Abschließend nochmals zwei Zitate des oben genannten Autors (2) (Übersetzung G. Mair):
"Wir sind eine evolutionäre Chimäre, intelligente Wesen, deren Intelligenz von tierischem Instinkt gesteuert wird. Dies ist die Ursache, warum wir gedankenlos die Biosphäre demontieren, und mit ihr die Perspektiven unserer eigenen Existenz."
"Wir sind allein auf diesem Planeten, mit aller Vernunft und allem Verständnis, das wir aufbringen können, und daher alleinverantwortlich für unsere Aktionen als Spezies."
Quellen:
(1) R. Wrangham, "Catching Fire" (New York: Basic Books, 2009)
(2) E. O. Wilson, "The Social Conquest of Earth" (London: W.W.Norton, 2012)
(3) V. Balter et al., "Evidence for dietary change but not landscape use in South African early hominins", Nature 489: 558–560 (2012),
doi:10.1038/nature11349
(4) L. Aiello, P. Wheeler, "The Expensive-Tissue Hypothesis: The Brain and the Digestive System in Human and Primate Evolution",
Current Anthropology 36: 199-221 (1995)
(5) R. Dunbar, "The Social Brain Hypothesis", Evolutionary Anthropology Bd. 6, Heft 5: 178-190 (1998)
(6) J. Mitani et. al., "Lethal intergroup aggression leads to territorial expansion in wild chimpanzees",
Current Biology 20 (12): R507-R508 (2010)
(7) aus (2); dort: S. Bowles, "Did warfare among ancestral hunter-gatherers affect the evolution of human social behaviors", Schience 324: 1295 (2009)
(8) aus (2); dort: T. Wynn, "Hafted spears and the archaeology of mind", Proceedings of the National Academy of Sciences, USA 106(24): 9544-9545 (2009)
(9) aus (2); dort: A. Patel, "Music as a transformative technology of the mind" in "Music, Language, and the Brain" (Oxford: University of Oxford Press, 2008)
(10) Nebenrechnung: Auszug aus Afrika 100 000 v.u.Z. angenommen mit 100 Personen --> Population 10 000 v.u.Z. 10 Mio. Menschen
--> jährliche Wachstumsrate 1,00013
Bevölkerung 1900 (vor Erfindung des Stickstoffdüngers) 1,7 Mrd. --> jährliche Wachstumsrate 1,0005
Bevölkerung 2012 7.0 Mrd. --> jährliche Wachstumsrate 1,013.
(11) J. McKee, "Sparing Nature" (New Brunswick: Rutgers University Press, 2005 [Copyright 2003]); dort:
R. MacPhee, ed., "Extinctions in Near Time: Causes, Contexts, and Consequences" (New York: Kluwer Academic/Plenum Publischers, 1999)